


オゾン等大気汚染物質に対する植物の応答に関与する遺伝子とその機能の解明
Summary
国立環境研究所では、設立以来オゾン等の大気汚染物質の植物影響に関する研究を行ってきました。特に、ガス暴露チャンバーを用いた実験により、植物のオゾン応答に関わる遺伝子とその機能を解明するための研究に取り組んできました。これらの研究で得られた成果は、オゾンのみならず様々なストレス因子に対する植物の応答への理解にもつながってきています。
オゾンなど大気汚染物質応答に関与する遺伝子の単離と遺伝子操作による耐性植物の作出
私たちが国立公害研究所(現・国立環境研究所)に着任した時点(1985-1990年)では、それまでに行われた生理・生化学的研究により、オゾン等のガス状大気汚染物質によって植物に種々の障害が生じ、それには活性酸素やエチレンなどの物質の関与が示唆されていました。しかし、それらの反応に関与する遺伝子は何ひとつ明らかにされておらず、障害を防ぐための有効な手法も確立されていませんでした。そこで私たちは、まず活性酸素の消去系に関連する酵素の遺伝子と、エチレン生成に関連する酵素の遺伝子を植物から単離し、その構造を決定するとともに、それらの遺伝子の発現がオゾンや二酸化硫黄により誘導されることを明らかにしました。さらに、それらの遺伝子を操作することにより、オゾンや二酸化硫黄に強い植物(タバコ)を作出することに成功しました。これらの成果は、活性酸素やエチレンが植物のオゾン障害に関与するという従来の仮説を裏付けるとともに、植物育種等の応用面にもつながるものとなりました。
植物のオゾン感受性への気孔開閉制御の関与の証明
2000年頃から国内外でシロイヌナズナやイネ等のモデル植物を中心に、網羅的に遺伝子を解読するゲノム解析が進み、その活用によりこれまで困難であった生理反応の解明を遺伝子のレベルから効率良く行えるようになってきました。そこで私たちもその流れに従って、シロイヌナズナの突然変異体を用いた分子遺伝学的研究に取り組みコラム4の図4参照、オゾン応答に関してこれまでに提案されていた仮説について、さらに裏付ける証拠やまだ知られていなかった新たな反応の関与を示唆する結果を次々と得ることに成功しました。その1つが気孔開閉の制御に関するものでした。
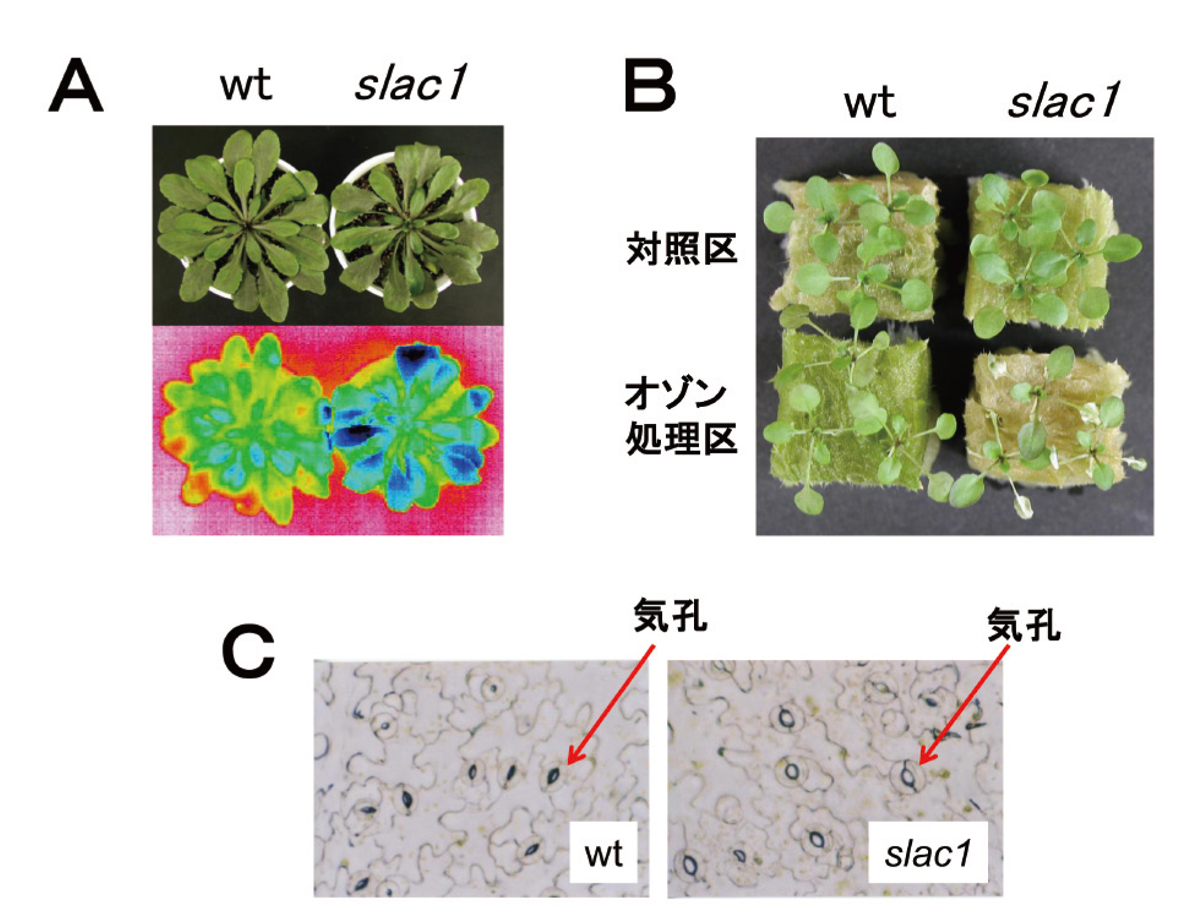
オゾンは葉の表面にある気孔を通って植物体内に吸収されることから、気孔開閉の制御がオゾンに対する最初の防御機構として重要であると考えられていました。しかし、それをはっきりと示す証拠はまだ得られていませんでした。シロイヌナズナのオゾン感受性突然変異体をいくつか単離してその性質を調べたところ、気孔反応に異常を示すものがあることを発見しました。この変異体は野生型と比べて気孔開度が大きく(図6A, C)、ガス状汚染物質のオゾン(図6B)や二酸化硫黄の他、乾燥にも高い感受性を示しました。これにより、植物のオゾン感受性への気孔開閉制御の関与がはっきりと示されました。さらに研究を進めた結果、この変異体の原因が、細胞膜上で物質の移動を制御するようなタンパク質の遺伝子が破壊され、機能しなくなったことによることがわかりました。
その後、ヘルシンキ大学のグループと九州大学のグループにより同様の変異体の単離と解析が行われ、この遺伝子(SLAC1と名付けられました)が、オゾン、乾燥、高濃度二酸化炭素条件下などにおいて、気孔が閉じる際に活性化される陰イオンチャンネルのものであることが示されました。さらにこれらの発見を契機として、気孔閉鎖メカニズムの研究が大きく進展し、今では、種々のシグナル伝達を介して気孔を取り囲む細胞膜上に存在するSLAC1やその他のタンパク質が次々に活性化されることで、気孔が閉じることが明らかになっています。
植物のオゾン応答における活性酸素の役割の詳細な解析
オゾンはそれ自身が反応性に富む活性酸素の一種であり、植物体内に吸収されると葉の組織中の水に溶け込み、細胞膜やその外側(アポプラスト)に存在する様々な物質を酸化すると考えられます。その初期反応に加え、種々の活性酸素種(ROS)が二次的に発生し、オゾン障害に関わることが示されてきました。中でも重要なのが、原形質膜上に存在するNADPHオキシダーゼ等によってアポプラストで産生されるROSと光照射下の葉緑体で光合成に伴って発生するROSです。前者は病原抵抗性反応の研究により見つけられ、病原体感染を知覚した葉の細胞が種々のシグナルを介してNADPHオキシダーゼ等を活性化し、アポプラストにおいてROSを自ら生成します。このようなROS生成は、病原体感染時のみならず種々のストレス条件下で起こり、オゾンストレス下でも障害の発生に関与することが、欧米の研究グループにより示されてきました。
一方、後者の光合成に伴って発生するROS生成は、オゾン障害が光照射下で生じやすいことから推論されてきましたが、最近、私たちの解析したシロイヌナズナのオゾン感受性突然変異体(オゾン耐性の低い突然変異体)の原因遺伝子が光呼吸という代謝系の酵素のものであることがわかり、これを裏付ける証拠となりました。この突然変異体は、光が強い場合にのみ高いオゾン感受性を示します(図7A)。光呼吸は、強光下において光合成の電子伝達系により葉緑体内で作られるエネルギー物質(NADPHとATP)を消費することにより、電子伝達をスムーズに進行させ、葉緑体内でのROS生成を抑える方向に作用すると考えられています。この光呼吸系の突然変異体が強光下でオゾンに高い感受性を示すことから、オゾンによるアポプラストにおけるROS生成と強光下の葉緑体内におけるROS生成が同時に起こると細胞死が強く誘導され、葉の可視障害が生じることが示唆されました(図7B)。オゾン以外にも種々のストレス因子によりアポプラストでROSが生成することや、強光下でストレスの程度が増すことが知られていることから、この仮説はオゾンのみならず他の多くのストレス応答にも適用できると考えられます。
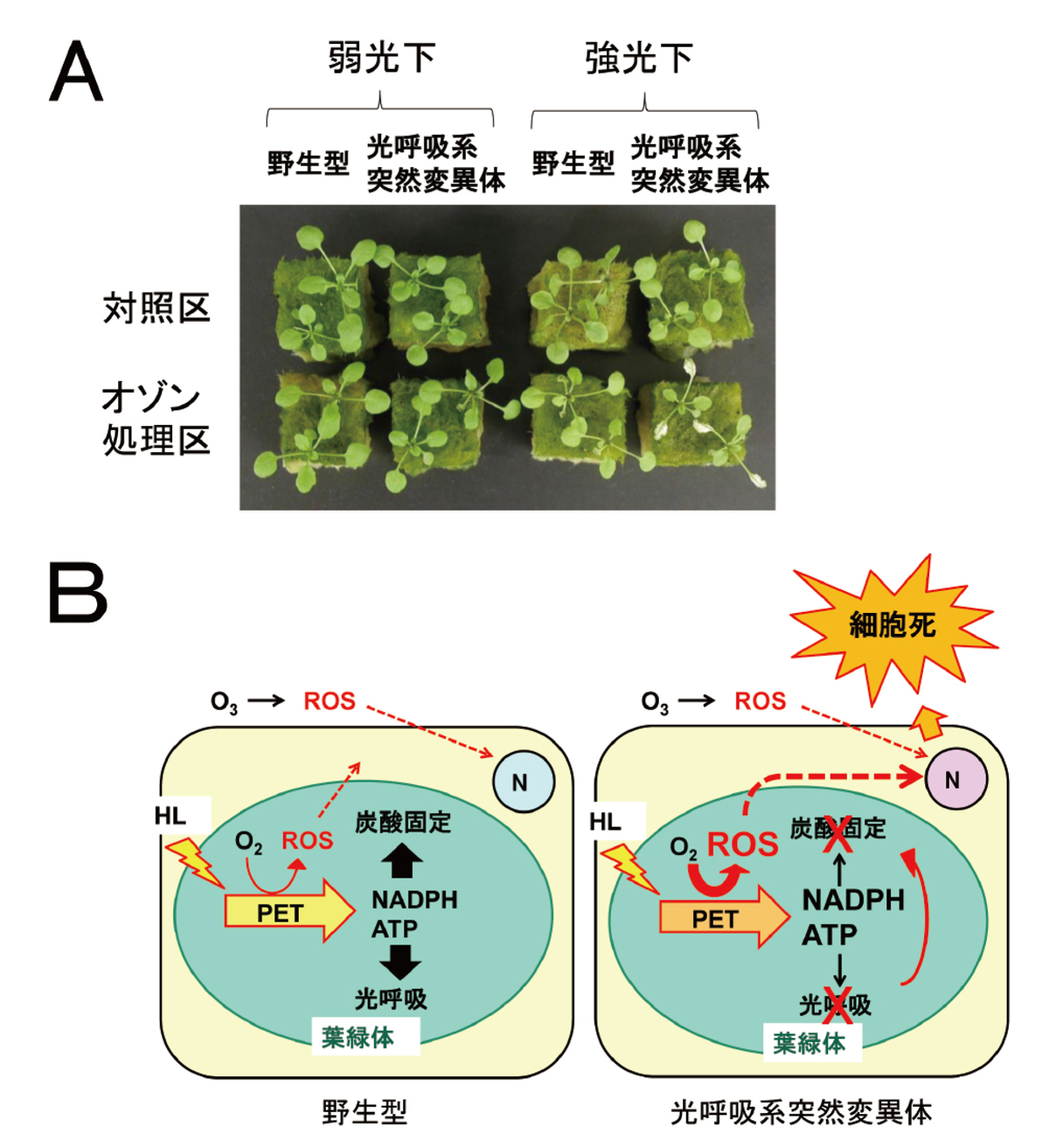
B. 光呼吸に異常のある突然変異体がオゾン感受性になる理由についての仮説。野生型の植物では(左図)、強光下において光合成の電子伝達系により葉緑体内で作られるエネルギー物質(NADPHとATP)が炭酸固定や光呼吸により消費され、過剰に蓄積されることはありません。ところが、光呼吸系突然変異体では(右図)、光呼吸や炭酸固定(カルビン回路の基質枯渇による)が阻害されるため、これらのエネルギー物質が十分消費されずに過剰蓄積します。その結果、電子伝達の流れが悪くなり、電子が酸素に受け渡されて活性酸素が多く発生します。この葉緑体における活性酸素生成とオゾンによる細胞外での活性酸素生成が同時に起こると、遺伝的プログラムにより細胞死が誘導されます。したがって、光呼吸は、強光下において、オゾン等のストレス因子に対する植物の耐性に大変重要な役割を担っていると考えられます。
HL:強光、N:核、O3:オゾン、PET:光合成電子伝達系、ROS:活性酸素







目次
関連新着情報
-
2022年12月23日
 植物にオゾン耐性を付与するタンパク質
植物にオゾン耐性を付与するタンパク質
~フィトシアニンによる新規ストレス防御機構の発見~(筑波研究学園都市記者会、環境省記者クラブ、環境記者会同時配付) -
2022年9月8日液体脂肪酸の光反応による活性酸素の生成
—海洋表面やエアロゾル界面の化学の理解に貢献— -
2021年4月22日我が国における金属由来の粒子酸化能の大気中濃度について、初めて予測に成功し、発生源別の寄与率を評価しました—健康影響の低減に効果的な大気汚染物質の削減に向けて—(筑波研究学園都市記者会、気象庁記者クラブ、自動車産業記者会、環境省記者クラブ、環境記者会、文部科学記者会、科学記者会、京都大学記者クラブ同時配布)
-
2020年6月30日エアロゾルのエイジングを研究する
大気中のエアロゾル粒子はどのように変質していくのか?
国立環境研究所「環境儀」第77号の刊行について(筑波研究学園都市記者会、環境記者会、環境省記者クラブ同時配付) -
2020年4月18日PM2.5濃度上昇が心停止の発生に影響?
~日本全国規模の人を対象とした疫学研究の成果~(文部科学記者会、科学記者会、厚生労働記者会、筑波研究学園都市記者会、環境省記者クラブ、環境記者会、他同時配付) -
2020年4月9日大気汚染物質を生成する「ホンモノ」と生成しない「ニセモノ」を見分ける~二次有機エアロゾル生成に関わるテルペン二量体を正確に検出~(筑波研究学園都市記者会、環境省記者クラブ、環境記者会、文部科学記者会、科学記者会同時配付)
-
2018年10月25日大気汚染物質のリスク評価手法に関するセミナー
—今後の有害大気汚染物質の健康リスク評価のあり方について—【終了しました】(筑波研究学園都市記者会、環境省記者クラブ、環境記者会同時配布) -
2018年1月12日「遺伝子から植物のストレスにせまる─オゾンに対する植物の応答機構の解明」
国立環境研究所「環境儀」第67号の刊行について(お知らせ)
(筑波研究学園都市記者会、環境記者会、環境省記者クラブ同時配付) -
2017年9月4日黄砂飛来の翌日に急性心筋梗塞が増える可能性
(文部科学記者会、科学記者会、熊本県内報道機関、筑波研究学園都市記者会、環境省記者クラブ、京都大学記者クラブ、大阪科学・大学記者クラブ同時配付) -
2017年8月1日
 環境GIS「大気汚染予測システム」をリニューアルしました
環境GIS「大気汚染予測システム」をリニューアルしました
-
2016年7月12日2015年のつくば大気質モニタリングデータを公開しました
-
2015年11月20日シベリアの森林火災によるPM2.5環境基準濃度レベルの超過について(お知らせ)
(筑波研究学園都市記者会、環境省記者クラブ同時配付) -
2015年10月13日環境GIS「大気汚染の常時監視結果」に2013年度データを追加しました
-
2015年8月19日環境数値データベース「大気環境データ」に
2013年度データを追加しました -
2015年6月12日日食を利用して太陽光が大気中のオゾンへ与える影響を調査
-
2015年4月30日光化学オキシダントでコメの収穫量が減る新たな要因を解明—イネの穂の枝分かれが関与—(筑波研究学園都市記者会、環境省記者クラブ同時配付)
-
2015年4月8日2014年のつくば大気質モニタリングデータを公開しました
-
2015年4月1日環境GIS「大気汚染予測システム」をリニューアルしました
-
2014年12月3日環境GIS「大気汚染の常時監視結果(2012年度)」を追加しました
-
2014年10月6日「環境と人々の健康との関わりを探る~環境疫学~」
国立環境研究所「環境儀」第54号の刊行について(お知らせ)
(筑波研究学園都市記者会、環境省記者クラブ同時配付)
関連記事
-
2021年10月29日
 温室効果ガスや大気汚染物質の排出量を迅速に把握する重要性とその方法特集 温室効果ガスや大気汚染物質の排出実態を迅速に把握する
温室効果ガスや大気汚染物質の排出量を迅速に把握する重要性とその方法特集 温室効果ガスや大気汚染物質の排出実態を迅速に把握する
-
2021年6月30日公害問題から地域の持続可能性に向けて【地域環境保全領域の紹介】
-
2016年8月31日気候変動対策と大気汚染対策の最適なバランスとは?特集 パリ協定とその先を見据えて
【研究ノート】 -
2013年10月31日大気汚染物質を含む環境汚染物質が免疫・アレルギーに及ぼす影響とその作用機構の解明特集 大気汚染の現状と健康影響評価
【シリーズ先導研究プログラムの紹介:『小児・次世代環境保健研究プログラム』から】 -
2013年10月31日大気汚染物質を細胞に直接曝露する気液界面細胞曝露装置特集 大気汚染の現状と健康影響評価
【研究ノート】 -
2011年6月30日環境健康研究センターの組織と研究プログラム【環境健康研究センターの紹介】
関連研究報告書
-
 2017年2月28日生物多様性研究プログラム(重点研究プログラム)
2017年2月28日生物多様性研究プログラム(重点研究プログラム)
平成23~27年度国立環境研究所研究プロジェクト報告 SR-116-2016 -
 2016年12月19日東アジア広域環境研究プログラム(重点研究プログラム)
2016年12月19日東アジア広域環境研究プログラム(重点研究プログラム)
平成23~27年度国立環境研究所研究プロジェクト報告 SR-115-2016 -
 2006年12月28日大陸規模広域大気汚染に関する国際共同研究(特別研究)
2006年12月28日大陸規模広域大気汚染に関する国際共同研究(特別研究)
平成13〜17年度国立環境研究所特別研究報告 SR-65-2006 -
 2006年12月28日大気中微小粒子状物質(PM2.5)・ディーゼル排気粒子(DEP)等の大気中粒子状物質の動態解明と影響評価プロジェクト(終了報告)
2006年12月28日大気中微小粒子状物質(PM2.5)・ディーゼル排気粒子(DEP)等の大気中粒子状物質の動態解明と影響評価プロジェクト(終了報告)
平成13〜17年度国立環境研究所特別研究報告 SR-74-2006 -
 2003年9月30日大気汚染・温暖化関連物質監視のためのフーリエ変換赤外分光計測技術の開発に関する研究(革新的環境監視計測技術先導研究)
2003年9月30日大気汚染・温暖化関連物質監視のためのフーリエ変換赤外分光計測技術の開発に関する研究(革新的環境監視計測技術先導研究)
平成12〜14年度国立環境研究所特別研究報告 SR-52-2003 -
 2001年6月25日大気有害化学物質監視用自動連続多成分同時計測センサー技術の開発に関する研究(革新的環境監視計測技術先導研究)
2001年6月25日大気有害化学物質監視用自動連続多成分同時計測センサー技術の開発に関する研究(革新的環境監視計測技術先導研究)
平成9〜11年度国立環境研究所特別研究報告 SR-39-2001